Features of the structure and function of the brain of various classes of vertebrates (fish, reptiles, birds, mammals). Brain (fish, amphibians, birds) Description of the structure of the brain of amphibians
Amphibians (amphibians).
Forebrain amphibians form two hemispheres, inside of which the lateral ventricles with choroid plexuses are located. In front of the forebrain lie the large olfactory bulbs. They are weakly delimited from the hemispheres and in tailless amphibians they grow together along the midline. Signals coming from the olfactory bulbs are analyzed in the forebrain, which is essentially the highest olfactory center. The roof of the forebrain is formed by the primary cerebral vault - the archipallium. It contains nerve fibers (white matter), and in the depths, underneath them, lie nerve cells. At the bottom of the forebrain there are clusters of neurons - striatal bodies.
Immediately behind the forebrain hemispheres is located diencephalon with a well-developed upper cerebral appendage - the pineal gland and a lower cerebral appendage - the pituitary gland. In general terms, the diencephalon is similar to that of fish.
The largest part of the brain in amphibians is midbrain. It looks like two hemispheres covered with bark. It includes the optic tract as a continuation of the optic nerves, and here the integration of the visual analyzer with other sensory pathways occurs and a center is formed that performs complex associative functions. Thus, the midbrain serves as the leading department of the central nervous system, where the information received is analyzed and response impulses are generated, i.e. Amphibians, like fish, have an ichthyopsid type of brain.
Rice. 7. The brain of a frog (dorsal view).
1 – forebrain hemispheres.
2 – olfactory lobes.
3 – olfactory nerves.
4 – diencephalon.
5 – midbrain.
6 – cerebellum.
7 – medulla oblongata.
8 – spinal cord.Cerebellum in most caudate and tailless amphibians, it is small in size and has the appearance of a transverse ridge at the anterior edge of the rhomboid fossa of the medulla oblongata. Poor development of the cerebellum reflects the unsophisticated motor coordination of amphibians. Most of the cerebellum consists of the median part (cerebellar body), where the integration of signals from muscle receptors and the vestibular system occurs.
In amphibians, like in fish, cerebellar nerve fibers are connected to the midbrain, brainstem and spinal cord. Vestibular-cerebellar connections determine the ability of animals to coordinate body movements.
Medulla oblongata in basic features similar to the medulla oblongata of fish. There are 10 pairs of cranial nerves originating from the brainstem.
Reptiles (reptiles).
Reptiles are true land animals that can live, reproduce and develop far from bodies of water. They belong to higher vertebrates. Their nervous system, due to their active and complex lifestyle, is better developed than that of amphibians.

Rice. 8. Lizard brain (according to Parker).
A - top view.
B – bottom view.
B – side view.
1 – forebrain; 2 – striatum; 3 – midbrain; 4 – cerebellum; 5 - medulla oblongata; 6 – funnel; 7 – pituitary gland; 8 - chiasm; 9 – olfactory lobes; 10 – pituitary gland; II – XII- head nerves.Forebrain significantly larger in size than amphibians and has a more complex structure; their ability to form conditioned reflexes increases, new connections with the external environment are established more quickly, and they can adapt to environmental changes better than their ancestors. The forebrain consists of two hemispheres, which, growing backward, cover the diencephalon with the exception of the pineal gland and the parietal organ. The enlargement of the forebrain occurs mainly due to the striatum (clusters of neurons) located in the area of the bottom of the lateral ventricles. They act as a higher integrative center, providing analysis of information entering the forebrain and the development of responses. Thus, it ceases to be only an olfactory center. This type of brain is called sauropsid. As for the brain vault, important transformations occur in it. In both hemispheres of the roof of the forebrain, for the first time in evolution, two islands of gray matter (cortical rudiments) appear - one of them is located on the medial and the other on the lateral side of the hemispheres. Only the medial insula, which represents the highest olfactory center, is functionally significant. In general, the islands of the cortex have a primitive structure and are called ancient cortex (archicortex). Most authors consider the islands of the cortex to be single-layered, although in crocodiles two or even three layers can be distinguished.
The olfactory lobes associated with the forebrain are well developed. In some species they occupy a sessile position, but more often they are differentiated into a bulb and a stem.
The study of the reptile forebrain is important for evolutionary neurohistology, because. they are a key point in the evolution of vertebrates, from which the development of the forebrain proceeded in two fundamentally different directions: along the striatal path with the predominant development of subcortical structures in birds and along the cortical path with the predominant development of cortical structures in mammals.
Diencephalon on a thin roof it has two bubble-like formations, one of which is located in front and is called the parietal, or parapineal, organ, and the second behind is the epiphysis (pineal gland). The parapineal organ performs a photosensitive function, and therefore it is also called the parietal eye. In fact, the parapineal organ and the pineal gland form a tandem, which is a regulator of the daily activity of animals. However, the parietal organ is not found in all reptiles. In such cases, the mechanism for regulating daily activity will be different: information about the length of daylight hours comes not from the parapineal organ, but from the visual system.
Midbrain represented by the colliculus and, in its main features, has the same organization that is characteristic of amphibians, however, reptiles are characterized by a more precise topographic representation of each of the sensory systems in the midbrain. In addition, almost all motor nuclei of the cerebellum and medulla oblongata interact with neurons in the roof of the midbrain. At the same time, the midbrain is losing its significance as the main integrative department of the central nervous system. This function passes to the forebrain.
Some of the visual and auditory fibers are sent along bypass pathways, bypassing the midbrain, to the forebrain. In the midbrain, the centers for ensuring automatic innate reactions of the body, obtained in the early stages of the evolution of vertebrates, are preserved. New forebrain centers take over the functions of the thecum and form new motor pathways.
Cerebellum due to the development of walking and running by reptiles, it is better developed than in amphibians. It consists of a central axial part, called the worm, and some have lateral lobes. The cerebellum is characterized by numerous connections with other parts of the nervous system related to locomotion. The spinocerebellar tract related here, present in both fish and amphibians, also extends to the dorsal part of the vermis. There are cerebellar connections with the vestibular apparatus, midbrain and rhombencephalon. Vestibular-cerebellar connections control the position of the body in space, and thalamic connections regulate muscle tone.
Medulla oblongata forms a sharp bend in the vertical plane, characteristic in general for amniotes.
There are 12 pairs of cranial nerves originating from the brainstem.
Birds.
Birds are a highly specialized group of vertebrates adapted to flight. Being physiologically close to reptiles, birds have a brain in many ways similar to that of reptiles, although they differ in a number of progressive features, which led to more complex and varied behavior. Their brain is quite large. If in reptiles its mass is approximately equal to the mass of the spinal cord, then in birds it is always larger. The brain bends are sharply expressed.
The enlargement of the brain is primarily due to the development forebrain, which essentially covers all parts of the brain with the exception of the cerebellum. Due to this, the midbrain is not visible from above, although it is well developed. The roof of the forebrain remains poorly developed. The cortex not only does not receive further development, but in the hemispheres the lateral island of the cortex disappears, while the medial one retains the significance of the highest olfactory center.
The increase in the size of the forebrain occurs due to the bottom, where the large striatal bodies, which are the leading part of the brain, are located. That is, birds retain sauropsid type of brain.
The olfactory lobes are very small, which is associated with the poor development of the sense of smell, and are closely adjacent to the forebrain.
Diencephalon small. In most birds, only the pineal gland is usually preserved, and the parapineal organ disappears in the late embryonic period. Information about the length of daylight hours does not come from the parapineal organ, but directly from the visual system. In the thalamus, the dorsal section is the most developed, which is the switching center of afferent connections with the forebrain. It contains a complex of nuclei on the path of descending fibers from the motor centers of the forebrain. Below it is the hypothalamus, connected to the pituitary gland, which is well developed in birds. The hypothalamus plays a major role in hormonal regulation of the body, maintaining homeostasis, sexual and eating behavior.
Midbrain consists of two large mounds. It contains the highest centers for processing visual and auditory information, as well as centers for regulating species-specific inherited forms of coordinated reactions, which are the basis of life. There are also sensory nuclei that perform an associative function, sending signals to the diencephalon and forebrain. At the same time, there is a decrease in the number of descending connections between the roof of the midbrain and motor centers due to the appearance in birds of striatal-reticular relatively autonomous connections between the striatal bodies of the forebrain and the reticular formation of the brain stem.
Cerebellum large and its structure becomes more complex. In front it comes into contact with the posterior edges of the forebrain hemispheres, and behind it covers a significant part of the medulla oblongata. The cerebellum is divided into a middle part (vermis) and lateral projections. Its worm is dotted with characteristic transverse grooves. The relatively complex structure of the cerebellum is due to complex movements that require high coordination during flight.Medulla oblongata relatively small, its lower side, even more than in reptiles, forms a downward bend, and in the region of the diencephalon there is an upward bend.
The cranial nerves of birds are represented in 12 pairs.
Mammals.
Mammals are the most highly organized class of vertebrates with a highly developed central nervous system. In this regard, the adaptive reactions of mammals to environmental conditions are complex and very advanced.
Forebrain large, it is significantly larger than all other parts of the brain. Its hemispheres grow in all directions, hiding the diencephalon. The midbrain is visible from the outside only in aplacentals and lower placentals, and in ungulates, carnivores, cetaceans and primates it is covered by the posterior part of the cerebral hemispheres. In anthropoids and humans, the occipital lobes of the forebrain are also pushed onto the cerebellum.
If initially, during evolution, the bulk of the telencephalon consisted of the olfactory lobes, then in mammals only the lower ones have developed olfactory lobes, and in the higher ones the olfactory lobes have the form of small appendages, divided into the olfactory bulb and the olfactory tract.
The increase in the relative size of the forebrain of mammals is associated primarily with the growth of its roof, and not the striatum, as in birds. The medullary vault (roof) is formed by gray matter called the cortex. The latter is a complex consisting of an ancient cloak (paleopalium), an old cloak (archipallium) and a new cloak (neopalium). The new cloak occupies an intermediate position, located between the old and ancient cloaks. The old mantle, or old cortex, is located medially and in the past it was called the hippocampus or the horn of Ammon. The ancient cloak, or ancient bark, occupies a lateral position.
The new cloak is usually called the neocortex (new cortex) and it is from this that the forebrain hemispheres are mainly composed. In this case, the surface of the hemispheres can be smooth (lisencephalic) or folded (with grooves and convolutions). In addition, regardless of this, from 4 to 5 lobes are distinguished in the hemispheres. The principle of dividing the forebrain into lobes is based on the topography of certain grooves and convolutions. The division into lobes in the lisencephalic (smooth) brain is conditional. Usually there are parietal lobes, temporal, occipital and frontal, and in higher primates and humans there is also a fifth lobe, which is called the insula. It is formed in the embryonic period due to the growth of the temporal lobe to the ventral side of the hemispheres.Taking the lisencephalic brain as the initial type of the cerebral hemispheres, three options for the development of the pattern of grooves are distinguished: longitudinal, arcuate and “primate type”. In the primate type, the groove in the frontal lobes is directed rostrally, and in the temporal lobes - ventro-dorsally
The location of the sulci and gyri can be significantly influenced by the shape of the brain. In most mammals, the brain is elongated in a rostro-caudal direction. However, in many dolphins the brain is expanded laterally and relatively shortened in length.
To characterize the forebrain of mammals, in addition to the grooves and convolutions, the nature of the distribution of neurons in the cortex (cytoarchitecture) is of great importance. The mammalian neocortex has a six-layer structure and is characterized by the presence of pyramidal cells, which are absent in the brains of other vertebrates. Especially large pyramidal cells (Betz cells) are located in the motor cortex. Their axons transmit nerve impulses to motor neurons of the spinal cord and motor neurons of the motor nuclei of the cranial nerves.
Different areas of the cerebral cortex are specialized areas for processing information coming from various sensory organs. There are sensory and motor areas. The latter form the descending pathways of nerve fibers to the brain stem and spinal motor nuclei. Between the sensitive and motor areas of the cortex there are integrative areas that combine the inputs of the sensory and motor areas of the cortex and determine the performance of specialized species-specific functions. In addition, there are associative zones of the cortex that are not associated with specific analyzers. They represent a superstructure over the rest of the cortex, providing thought processes and storage of specific and individual memory.
The entire complex of zones distributed in the cortex is associated with functional specialization of fields. In this case, the morphological and functional boundaries of the fields coincide quite accurately. The criterion for identifying a particular field is a change in the distribution of cellular elements in the cortex or the emergence of a new sublayer in it.
Features of the architectonics of certain fields are a morphological expression of their functional specialization. The reason for the change in cytoarchitecture in the fields is an increase in the number of ascending and descending nerve fibers. Topological maps of fields have now been created for humans and for many laboratory animals.
The fields of the cerebral cortex are part of certain lobes and are themselves divided into functional zones associated with specific organs or their parts and have an ordered internal structure. In each field or zone there are so-called modules vertical orderliness of the organization of the cortex. The module has either the form of a column or a glomerulus, which includes neurons located throughout the thickness of the cortex. The column contains a group of 110 neurons located between a pair of capillaries running across the diameter of the cortex.
At the stage of formation of the brain of the most ancient hominids, the area where the action of natural selection was directed was the cortex and, first of all, its following sections: the inferior parietal, inferior frontal and temporo-parietal regions. The survival advantage was given to those individuals, and then to those populations of emerging people who turned out to be advanced in terms of the development of some elements of the parts of the cortex (larger area of fields, more diverse and mobile connections, improved conditions of blood circulation, etc.). the development of new connections and structures in the cortex provided new opportunities for the manufacture of tools and team building. In turn, a new level of technology, the beginnings of culture and art through natural selection contributed to the development of the brain.
To date, an idea has been formed about a specific systemic complex of the human forebrain cortex, including the inferior parietal, posterior superior temporal and inferior frontal lobes of the cortex. This complex is associated with higher functions - speech, work activity and abstract thinking. In general, it is the morphological substrate of the second signaling system. This system does not have its own peripheral receptors, but uses the old receptor apparatus of various sense organs. For example, it has been established that the tongue has a special part of the tactile apparatus, the development of which determines the sequence of sound production in the initial stages of the formation of a child’s articulate speech.
The submantle structures of the forebrain include the basal ganglia, the striatum (ancient, old and new) and the septal field.
In various parts of the forebrain and diencephalon there is a complex of morphofunctional structures called limbic system. The latter has numerous connections with the neocortex and the autonomic nervous system. It integrates brain functions such as emotions and memory. Removing part of the limbic system leads to emotional passivity of the animal, and its stimulation leads to hyperactivity. The most important function of the limbic system is interaction with memory mechanisms. Short-term memory is associated with the hippocampus, and long-term memory is associated with the neocortex. Through the limbic system, the individual experience of the animal is extracted from the neocortex, the motor control of internal organs, and hormonal stimulation of the animal occur. Moreover, the lower the level of development of the neocortex, the more the animal’s behavior depends on the limbic system, which leads to the dominance of emotional-hormonal control over decision making.
In mammals, descending connections from the neocortex to the limbic system enable the integration of a wide variety of sensory inputs.
With the appearance of the first rudiments of the cortex in reptiles, a small bundle of nerve fibers connecting the left and right hemispheres separated from the mantle commissure. In placental mammals, such a bundle of fibers is much more developed and is called the corpus collosum. The latter provides the function of interhemispheric communications.
Diencephalon, like other vertebrates, consists of the epithalamus, thalamus and hypothalamus.
The development of the neocortex in mammals led to a sharp increase in the thalamus, and, above all, the dorsal one. The thalamus contains about 40 nuclei, in which ascending pathways switch to the last neurons, the axons of which reach the cerebral cortex, where information coming from all sensory systems is processed. At the same time, the anterior and lateral nuclei process and transmit visual, auditory, tactile, gustatory and interoceptive signals to the corresponding projection zones of the cortex. There is an opinion that pain sensitivity is not projected into the forebrain cortex, and its central mechanisms are located in the thalamus. This assumption is based on the fact that irritation of different areas of the cortex does not cause pain, while irritation of the thalamus causes severe pain. Some of the nuclei of the thalamus are switching, and the other part are associative (from them there are paths to the associative zones of the cortex). In the medial part of the thalamus there are nuclei that, with low-frequency electrical stimulation, cause the development of inhibitory processes in the cerebral cortex, leading to sleep. High-frequency stimulation of these nuclei causes partial activation of cortical mechanisms. Thus, the thalamocortical regulatory system, by controlling the flow of ascending impulses, is involved in organizing the change between sleep and wakefulness.
If in lower vertebrates the higher sensory and associative centers are located in the midbrain, and the dorsal thalamus is a modest integrator between the midbrain and the olfactory system, then in mammals it is the most important center for switching auditory and somatosensory signals. At the same time, the somatosensory area has become the most prominent formation of the diencephalon and plays a huge role in the coordination of movements.
It should be noted that the complex of thalamic nuclei is formed both due to the primordium of the diencephalon and due to migration from the midbrain.
The hypothalamus forms developed lateral protrusions and a hollow stalk - a funnel. The latter ends in the posterior direction with the neurohypophysis tightly connected to the adenohypophysis.
The hypothalamus is the highest center for regulating endocrine functions of the body. It combines endocrine regulatory mechanisms with nervous ones. In addition, it is the highest center of the sympathetic and parasympathetic divisions of the autonomic nervous system.
The epithalamus serves as a neurohumoral regulator of daily and seasonal activity, which is combined with the control of puberty in animals.
Midbrain forms a quadrigeminal region, the anterior tubercles of which are connected with the visual analyzer, and the posterior ones with the auditory one. By the ratio of the relative sizes of the anterior and posterior tubercles, one can judge which of the systems, auditory or visual, is predominant. If the anterior tubercles are better developed, this means visual afferentation (ungulates, many predators and primates), if the posterior ones, then auditory afferentation (dolphins, bats, etc.).
The tagment is divided into sensory and motor zones. The motor zone contains the motor nuclei of the cranial nerves and the descending and ascending spinocerebral fibers.
In connection with the development of the neocortex in mammals as a higher integrative center, the innate reactions of the midbrain allowed the cortex to “not engage” in primitive forms of species-specific reactions to external signals, while complex associative functions were taken over by specialized fields of the cortex.
Cerebellum in mammals it acquires the most complex structure. Anatomically, it can be divided into a middle part - the vermis, hemispheres located on both sides of it and flocculonodular lobes. The latter represent the phylogenetically ancient part - Archicerebellum. The hemispheres are in turn divided into anterior and posterior lobes. The anterior lobes of the hemispheres and the posterior part of the cerebellar vermis represent the phylogenetically old cerebellum - paleocerebellum. Phylogenetically, the youngest part of the cerebellum, the neocerebellum, includes the anterior part of the posterior lobes of the cerebellar hemispheres.
The cerebellar hemispheres are divided into the upper surface, which forms the cerebellar cortex, and clusters of nerve cells - the cerebellar nuclei. The cerebellar cortex is built according to a single principle and consists of 3 layers. The cerebellum is connected to other parts of the central nervous system by three pairs of peduncles formed by bundles of nerve fibers. The hind legs consist primarily of proprioceptive fibers that come from the spinal cord. The middle crus are composed of fibers connecting the cerebellum and the forebrain cortex, and the anterior crura are formed by descending fibers connecting the cerebellum and midbrain.Vestibulocerebellar connections determine the ability of animals to coordinate body movements, which is the main function of the archicerebellum. In addition, new, more powerful cerebellar pathways have formed in mammals due to the emergence of the dentate nucleus of the cerebellum. It receives fibers from various parts of the cerebellar hemispheres and transmits signals to the thalamus, where sensorimotor signals are integrated with the activity of the cortical centers of the forebrain. Book
... functional phylogeny morpho-functional... central nervous, endocrine, respiratory and others systemsFunctional state...
From the fundamental sciences of morphology, physiology, physics, evolutionary teaching - to the ecology of intelligent systems, medicine, public health (1)
Book... functional, parametric, synergetic characteristics phylogeny and ontogenesis of the human brain. The body of knowledge about morpho-functional... central nervous, endocrine, respiratory and others systemsFunctional state...
From the fundamental sciences of morphology, physiology, physics, evolutionary teaching - to the ecology of intelligent systems, medicine, public health (2)
Book... functional, parametric, synergetic characteristics phylogeny and ontogenesis of the human brain. The body of knowledge about morpho-functional... central nervous, endocrine, respiratory and others systemsFunctional state...
Physiology of the central nervous system (1)
DocumentCoding - transformation information in... invertebrates and lower vertebrates animals... connection nervous And endocrinesystems. ... functional systems that receive signals from organsfeelings after preliminary processing in the central nervous ...
Boris Guryevich Meshcheryakov Vladimir Petrovich Zinchenko Large psychological dictionary Contents
Document... organsfeelings(and the human body in general) in combination with functional asymmetry... nervous cells ( nervous G.) outside the c. n. With. U vertebrates animals G. are located along the spinal cord and in the walls of the internal organs. U invertebrates ...
, reptiles (reptiles), birds, their nests, eggs and voices, and mammals (animals) and traces of their vital activity,
20
colored laminated definition tables, including: aquatic invertebrates, diurnal butterflies, fish, amphibians and reptiles, wintering birds, migratory birds, mammals and their tracks,
4
pocket field determinant, including: inhabitants of reservoirs, birds of the middle zone and animals and their traces, as well as
65
methodological benefits And 40
educational and methodological films By methods carrying out research work in nature (in the field).
Amphibian Anatomy: An Overview
Body structure or anatomy.
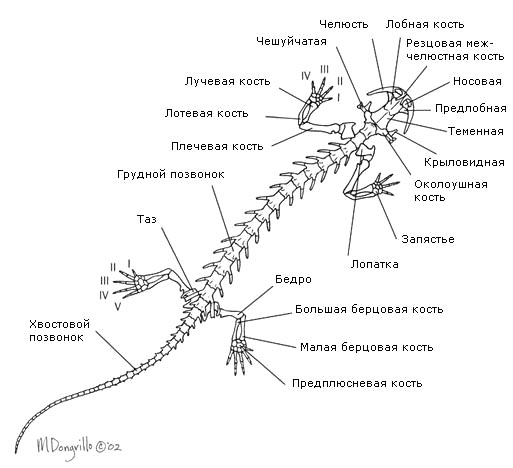
Body divided into head, torso, tail (only in caudates and legless animals) and limbs, which may be absent. Head mobile, connected to the body. The skeleton and spine are divided into departments. Ribs, if any, are attached to the trunk vertebrae.
Amphibians have two pairs of primary five-toed fingers limbs; the rudimentary forms of the neck provide them with the ability to move their heads independently.
Leather naked, devoid of scales. Epidermis rich in multicellular glands, which provide the presence of a liquid film on the surface of the skin, without which gas exchange is impossible during skin respiration. The epidermis is multilayered, the corium is thin, but richly saturated with capillaries.

In the lower layers of the epidermis and in the corium are located pigment cells, causing species-specific coloring.
Limb skeleton formed by the skeleton of the limb girdle and the skeleton of the free limbs.
Shoulder girdle lies in the thickness of the muscles and includes paired shoulder blades, collarbones and crow bones connected to the sternum. Skeleton forelimb consists of the shoulder (humerus), forearm (radius and ulna bones) and hand (bones of the wrist, metacarpus and phalanges of the fingers).
Pelvic girdle consists of paired iliac ischial and pubic bones fused together. It is attached to the sacral vertebra through the ilia. Included in the skeleton hind limb includes the thigh, lower leg (tibia and fibula) and foot. Bones of the tarsus, metatarsus and phalanges of the fingers. The sacrum consists of only one vertebra.
Propulsion system.
The movement pattern of amphibians is quite monotonous and can be reduced to two main types.
Fossil and modern tailed amphibians have retained their characteristic fish the main type of movement is with the help of strong lateral bends of the whole body, but with support on short legs when moving on the ground. With short limbs, the lateral bends of the body increase the length of the step, and the bends of the tail help maintain balance. When moving in water, the limbs do not play any noticeable role. Legless animals also move with the help of the bends of the whole body.
Tailless amphibians move on land jumping, lifting the body into the air with a sharp push of both hind limbs. Short-legged species, such as toads, in addition to jumping, can slowly step, sequentially rearranging the limbs.
Tailless in the water swim, vigorously working with the hind limbs (breaststroke style, but without the participation of the forelimbs). It is believed that powerful hind limbs developed as an adaptation to swimming, and only later were used for jumping on land.
Amphibians have a rather large, wide head, which goes directly into wide and short body. The frontal and parietal bones are fused into the paired frontoparietal bone. IN skull It is characteristic that the maxillo-palatine apparatus and the quadrate bone are fixedly connected to the skull; the two condyles of the skull belong to the first cervical vertebra that actually merged with it, so that the first vertebra of amphibians is essentially the second.
Brain amphibians differ from the fish brain in the greater development of the anterior section ( forebrain), containing a large number of nerve cells (gray matter). Hemispheres the forebrain are small and completely divided. The parts of the brain lie in the same horizontal plane. Olfactory lobes are highly developed. Cerebellum very poorly developed due to low mobility and monotonous nature of movements. There are 10 pairs of cranial nerves. Larvae have organs side line.
Spinal cord better developed than the head. The brain consists of 5 departments: forebrain, diencephalon, medulla oblongata, middle brain, cerebellum. Intermediate the brain is well developed. Oblong The brain is the center of the respiratory, circulatory and digestive systems. Average the brain is relatively small.
Organs of touch well developed. Organs side line signal amphibians about wave-like fluctuations in water. They are given to them for active location of the water space, especially in muddy water or at night, and completely replace vision. Being organs of remote touch, such living devices also sense vibrations caused by the movements of underwater inhabitants. The organs of the lateral lines are located on the surface of the skin of amphibians that live exclusively in water, and each species has its own characteristics.
The whole organ of touch is leather, which contains tactile nerve endings.
The mouth also contains organs of touch in the form of taste buds. Teeth may or may not be present in some species. The teeth, like those of reptiles, are adapted only for grasping and holding prey, but cannot serve to chew it. Sounds can only be made by tailless amphibians, and even then predominantly by males.
Nasal cavity equipped with posterior nasal openings and nasolacrimal ducts.
Eyes similar to the eyes of fish, but do not have a silvery shell, neither a reflective shell, nor a crescent-shaped process. Accommodation the eye is carried out by moving the lens. The eyes are adapted for long-distance vision. There are no lacrimal glands, but there is a Harderian gland, the secretion of which moistens the cornea and protects it from drying out. The cornea is convex. The lens has the shape of a biconvex lens, the diameter of which varies depending on the lighting; accommodation occurs due to changes in the distance of the lens to the retina. Many have developed color vision.

Structure ear differs in tailless and tailed amphibians.
Musculature divided into the muscles of the trunk and limbs. The trunk muscles are segmented. Groups of special muscles provide complex movements of lever limbs. The levator and depressor muscles are located on the head. Through contractions of muscles or muscle groups, amphibians can perform complex movements. The muscles of the limbs are especially well developed.
Digestive system amphibians have almost the same structure as fish. All amphibians feed only mobile prey. The tongue is located at the bottom of the oropharyngeal cavity. The ducts of the salivary glands open into the oropharyngeal cavity, the secretion of which does not contain digestive enzymes. From the oropharyngeal cavity, food enters the stomach through the esophagus, and from there into the duodenum. The ducts of the liver and pancreas open here. Digestion of food occurs in the stomach and duodenum. The small intestine passes into the large intestine, ending in the rectum, which forms an extension - the cloaca. Unlike fish, the hindgut does not open directly outward, but into a special extension called the cloaca. The ureters and excretory ducts of the reproductive organs also open into the cloaca.

Respiratory organs in amphibians are:
- lungs (special air breathing organs);
- skin and mucous lining of the oropharyngeal cavity (additional respiratory organs);
- gills (in some aquatic inhabitants and in tadpoles).
Most species (except lungless salamanders) have lungs small volume, in the form of thin-walled bags, braided with a dense network of blood vessels. Each lung opens with an independent opening into the laryngeal-tracheal cavity (the vocal cords are located here, opening a slit into the oropharyngeal cavity). Air is forced into the lungs by changing volume oropharyngeal cavity: air enters the oropharyngeal cavity through the nostrils when its bottom is lowered. When the bottom rises, air is pushed into the lungs.
Throat several times per second pulled down, due to which a rarefied space is created in the oral cavity. Then the air penetrates through the nostrils into the oral cavity, and from there into the lungs. It is pushed back under the action of the muscles of the body walls. An amphibian immersed in water completely switches to cutaneous respiration.
Circulatory system closed, consists of a large and small circle of blood circulation. The appearance of the second circle is associated with the acquisition of pulmonary breathing. The body has cutaneous pulmonary arteries (carry venous blood to the lungs and skin), carotid arteries (supply the organs of the head with arterial blood), and aortic arches carry mixed blood to the rest of the body organs.

I - venous sinus; II - right atrium; III - left atrium; IV - ventricle; V - arterial trunk;
1 - pulmonary cutaneous artery; 2 - aortic arch; 3 - carotid artery; 4 - lingual artery; 5 - carotid gland; 6 - subclavian artery; 7 - common aorta; 8 - intestinal artery; 9 - cutaneous artery; 10 - pulmonary vein; 11 - light; 12 - posterior vena cava; 13 - cutaneous vein; 14 - abdominal vein; 15 - liver; 16 - renal vein.
Pulmonary circulation- pulmonary, begins with the cutaneous pulmonary arteries, carrying blood to the respiratory organs (lungs and skin); From the lungs, oxygenated blood is collected in paired pulmonary veins, which flow into the left atrium.
Systemic circulation begins with the aortic arches and carotid arteries, which branch into organs and tissues. Venous blood enters the right atrium through the paired anterior vena cava and the unpaired posterior vena cava. In addition, oxidized blood enters the anterior vena cava, and therefore the blood in the right atrium is mixed. Since the body's organs are supplied with mixed blood, amphibians have a low metabolic rate and are therefore cold-blooded animals.
The aorta passes into the branchial arches and branches first in the external gills, and later in the internal ones. The blood flows back through a vein running along the tail, and then branches on the surface of the yolk sac and returns through the yolk veins back to the atrium. Later, the portal systems of the liver and kidneys gradually form. At the end of the larval stage, gill respiration is gradually replaced by pulmonary respiration; the anterior branchial arches turn into the cephalic arteries, and the middle ones form the aorta.
Heart three-chamber. It consists of two atria (in the right atrium the blood is mixed, mainly venous, and in the left - arterial) and one ventricle. Inside the walls of the ventricle, folds form that prevent the mixing of arterial and venous blood. An arterial cone, equipped with a spiral valve, emerges from the ventricle.
The right atrium receives venous blood, the left atrium receives arterial blood (from the lungs and skin). Venous and arterial blood only partially mix in the cavity of the ventricle, the walls of which have a complex system of muscular crossbars. Mainly venous blood is sent to the pulmonary veins, the aortic arches are filled with mixed blood, and only the carotid arteries receive arterial blood.
The heart is formed in the larvae very early and immediately begins to act. Initially it represents a simple bag, which is subsequently divided into separate parts.
Excretory organs- paired trunk kidneys, from which ureters depart, opening into the cloaca. In the wall of the cloaca there is an opening of the bladder into which urine that enters the cloaca from the ureters flows. There is no reabsorption of water in the trunk kidneys. After filling the bladder and contracting the muscles of its walls, concentrated urine is discharged into the cloaca and thrown out. Some metabolic products and a large amount of moisture are released through the skin. These features did not allow amphibians to completely transition to a terrestrial lifestyle. In larvae in the early stages of development, the so-called head kidney, or preference. Also, all amphibians have a lobed liver, gall bladder, and pancreas.
Reproductive system. All amphibians are dioecious. In most amphibians, fertilization external(in the water). During the breeding season, paired ovaries filled with mature eggs fill almost the entire abdominal cavity of females. Ripe eggs fall into the abdominal cavity of the body, enter the funnel of the oviduct and, after passing through it, are brought out through the cloaca. Males have paired testes. The seminiferous tubules extending from them enter the ureters, which at the same time serve as vas deferens for the males. They also open into the cloaca. The germ cells enter the cloaca through tubular ducts and are thrown out from there.
Muscular system. It differs from such fish mainly in the greater development of the muscles of the limbs and the greater differentiation of the trunk muscles, consisting of a complex system of individual muscles. As a result, the primary muscle segmentation is disrupted, although in some abdominal and dorsal muscles it still remains quite distinct.
Nervous system. The brain of amphibians differs from the brain of fish mainly in the greater development of the forebrain, the complete separation of its hemispheres and the underdeveloped cerebellum, which is only a small ridge of nerve substance covering the anterior part of the fourth ventricle. The development of the forebrain is expressed not only in its enlargement and differentiation, but also in the fact that, in addition to the bottom of the lateral ventricles, their sides and roof contain nerve matter, i.e. in amphibians a real brain vault appears - the archipallium (from modern fish the archipallium present in lungfishes). The olfactory lobes are only weakly delimited from the hemispheres. The diencephalon is only slightly covered from above by neighboring sections. The parietal organ is attached to its roof, and a well-defined funnel extends from the bottom, to which the pituitary gland is attached. Although the midbrain is a significant section, it is relatively smaller than that of fish. Underdevelopment of the cerebellum, as in lungfishes, is associated with the simplicity of body movements: amphibians are generally sedentary animals, but in those that, like frogs, can make rapid movements, they are limited to jumping, i.e., very simple movements. From the brain, like in bony fishes, only 10 pairs of head nerves depart; The XII pair (hypoglossal nerve) extends outside the cranium, and the XI pair (accessory nerve) is not developed at all.

. I - top; II - bottom; III - side; IV - in longitudinal section (according to Parker):
1 - forebrain hemispheres, 2 - olfactory lobe, 3 - olfactory nerve, 4 - diencephalon, 5 - optic chiasm, 6 -funnel, 7 - pituitary gland, 8 - midbrain, 9 - cerebellum, 10 - medulla oblongata, 11 - fourth ventricle, 12 - spinal cord, 13 - third ventricle, 14 - aqueduct of Sylvius,
III - X - head nerves, XII - hypoglossal nerve

, scheme (according to Gregory):
1 - cranium, 2 - medulla oblongata, 3 - auditory nerve, 4 - semicircular canals, 5 - middle ear cavity, 6 - eustachian tube, 7 - pharynx, 8 - eardrum, 9 - eardrum
The frog has 10 pairs of true spinal nerves. The three anterior pairs take part in the formation of the brachial plexus, which innervates the forelimbs, and the four posterior pairs take part in the formation of the lumbosacral plexus, which innervates the hind limbs.
The sympathetic nervous system of the frog, like that of all amphibians, is very well developed and is represented mainly by two nerve trunks that stretch on both sides of the spine and are formed by a chain of nerve ganglia connected to each other by cords and connected to the spinal nerves.
More interesting articles
Brain frogs, like other amphibians, are characterized by the following features compared to fish:
a) progressive development of the brain, expressed in the separation of the paired hemispheres by a longitudinal fissure and the development of the gray matter of the ancient cortex (archipallium) in the roof of the brain;
b) weak development of the cerebellum;
c) weak expression of the bends of the brain, due to which the intermediate and middle sections are clearly visible from above.
Diamond brain(rhombencephalon)
Medulla oblongata (myelencephalon, medulla oblongata), into which the spinal cord passes cranially, it differs from the latter in its greater width and the departure from its lateral surfaces of the large roots of the posterior cranial nerves. On the dorsal surface of the medulla oblongata there is diamond-shaped fossa (fossa rhomboidea), accommodating fourth cerebral ventricle (ventriculus quartus). On top it is covered with a thin vascular cap, which is removed along with the meninges. The ventral fissure, a continuation of the ventral fissure of the spinal cord, runs along the ventral surface of the medulla oblongata. The medulla oblongata contains two pairs of cords (bundles of fibers): the lower pair, separated by the ventral fissure, are motor, the upper pair are sensory. The medulla oblongata contains the centers of the maxillary and sublingual apparatus, the organ of hearing, as well as the digestive and respiratory systems.
Cerebellum located in front of the rhomboid fossa in the form of a high transverse ridge as an outgrowth of its anterior wall. The small size of the cerebellum is determined by the small and uniform mobility of amphibians - in fact, it consists of two small parts, closely connected with the acoustic centers of the medulla oblongata (these parts are preserved in mammals as fragments of the cerebellum (flocculi)). The body of the cerebellum - the center of coordination with other parts of the brain - is very poorly developed.
Midbrain(mesencephalon) when viewed from the dorsal side, it is represented by two typical optic lobes (lobus opticus s. tectum opticus), having the appearance of paired ovoid elevations forming the upper and lateral parts of the midbrain. The roof of the optic lobes is formed by gray matter - several layers of nerve cells. The tectum in amphibians is the most significant part of the brain. The optic lobes contain cavities that are lateral branches cerebral (Sylvii) aqueduct (aquaeductus cerebri (Sylvii), connecting the fourth cerebral ventricle with the third.
The bottom of the midbrain is formed by thick bundles of nerve fibers - cerebral peduncles (cruri cerebri), connecting the forebrain with the medulla oblongata and spinal cord.
Forebrain(prosencephalon) consists of the diencephalon and telencephalon, lying sequentially.
Diencephalon visible from above as a rhombus, with sharp angles directed to the sides.
Parts of the diencephalon lie around a vertically located wide fissure third cerebral ventricle (ventriculus tertius). Lateral thickening of the walls of the ventricle - visual cusps or thalamus. In fish and amphibians, the thalamus is of secondary importance (as coordinating sensory and motor centers). The membranous roof of the third cerebral ventricle - the epithalamus or epithalamus - does not contain neurons. It contains the superior medullary gland - pineal gland (epiphisis). In amphibians, the pineal gland already serves as a gland, but has not yet lost the features of the parietal organ of vision. In front of the epiphysis, the diencephalon is covered with a membranous roof, which orally turns inward and passes into the anterior choroid plexus (choroid tectum of the third ventricle), and then into the endplate of the diencephalon. Inferiorly the ventricle narrows, forming pituitary funnel (infundibulum), the inferior medullary gland is attached to it caudoventrally - pituitary gland (hypophisis). In front, on the border between the bottom of the terminal and intermediate parts of the brain, there is chiasma nervorum opticorum). In amphibians, most of the fibers of the optic nerves are not retained in the diencephalon, but go further to the roof of the midbrain.
Telencephalon its length is almost equal to the length of all other parts of the brain. It consists of two parts: the olfactory brain and two hemispheres, separated from each other sagittal (arrow-shaped) fissure (fissura sagittalis).
Hemispheres of the telencephalon (haemispherium cerebri) occupy the posterior two-thirds of the telencephalon and hang over the anterior part of the diencephalon, partially covering it. There are cavities inside the hemispheres - lateral cerebral ventricles (ventriculi lateralis), caudally communicating with the third ventricle. In the gray matter of the cerebral hemispheres of amphibians, three areas can be distinguished: dorsomedially there is the old cortex or hippocampus (archipallium, s. hippocampus), laterally - ancient bark(paleopallium) and ventrolaterally - the basal ganglia, corresponding striata (corpora striata) mammals. The striatum and, to a lesser extent, the hippocampus are correlative centers, the latter associated with olfactory function. The ancient cortex is an exclusively olfactory analyzer. On the ventral surface of the hemispheres, grooves are visible that separate the striatum from the ancient cortex.
Olfactory brain (rhinencephalon) occupies the anterior part of the telencephalon and forms olfactory lobes (bulbs) (lobus olfactorius), soldered in the middle with each other. They are separated from the hemispheres laterally by the marginal fossa. The olfactory lobes anteriorly contain the olfactory nerves.
10 pairs extend from the frog's brain cranial nerves. Their formation, branching and zone of innervation are not fundamentally different from those in mammals
137. Look at the pictures. Write the names of the frog's body parts. What organs are located on her head? Write their names.
138. Study the table “Class Amphibians. The structure of a frog." Look at the drawing. Write the names of the internal organs of the frog, indicated by numbers.

139. Describe the structure of the amphibian brain.
The brain of amphibians has more progressive features: larger forebrain sizes, complete separation of the hemispheres. The midbrain is relatively small. The cerebellum is small because amphibians have monotonous movements. There are 10 pairs of cranial nerves leaving the brain. Divisions of the brain: anterior, middle, cerebellum, medulla oblongata, intermediate.
140. Study the table “Class Amphibians. The structure of a frog." Look at the drawing. Label the parts of the frog's skeleton indicated by numbers.

1. skull
2. shoulder blade
3. shoulder
4. forearm
5. brush
6. foot
7. shin
8. thigh
9. urostyle
10. spine.
141. Look at the drawing. Write the names of the parts of the frog's digestive system indicated by numbers. How is digestion carried out in a frog?

All amphibians feed only on mobile prey. At the bottom of the oral cavity is the tongue. When catching insects, it is thrown out of the mouth, and the prey sticks to it. The upper jaw has teeth that serve only to hold prey. When swallowing, the eyeballs help push food into the esophagus from the oropharynx.
The ducts of the salivary glands open into the oropharynx, the secretion of which does not contain digestive enzymes. From the oropharynx, food enters the stomach through the esophagus, and from there into the duodenum. The ducts of the liver and pancreas open here. Digestion of food occurs in the stomach and duodenum. The small intestine passes into the rectum, which forms an extension - the cloaca.
142. Draw a diagram of the structure of a frog’s heart. Which blood is called arterial and which is called venous?
Arterial blood comes from the lungs and is rich in oxygen. Venous blood goes to the lungs.

143. Describe the process of reproduction and development of a frog. Indicate the similarities in the reproduction of amphibians and fish.
Amphibians breed in shallow, well-warmed areas of water bodies. The reproductive organs of males are the testes, and the reproductive organs of females are the ovaries. Fertilization is external.
Frog development:
1 - egg;
2 - tadpole at the moment of hatching;
3 - development of fin folds and external gills;
4 - stage of maximum development of external gills;
5 - stage of disappearance of external gills; 6 - stage of appearance of the hind limbs; 7 - stage of dismemberment and mobility of the hind limbs (the forelimbs are visible through the integument);
8 - stage of release of the forelimbs, metamorphosis of the oral apparatus and the beginning of tail resorption;
9 - stage of landfall.
144. Fill out the table.
The structure and significance of the frog's sense organs.

145. Complete laboratory work “Features of the external structure of a frog in connection with its lifestyle.”
1. Consider the features of the external structure of the frog. Describe the shape of its body, the color of its back and abdomen.
The frog's body is divided into head, torso and limbs. Long hind legs with webbed toes allow it to jump on land and swim in water. On the flattened head of the frog there is a large mouth slit, large bulging eyes located on elevations and a pair of nostrils. On the sides of the head behind the eyes are rounded eardrums (eardrums). The frog's eyes are large and bulging. The eyes are equipped with movable eyelids. Male green frogs have resonators, or vocal sacs, in the corners of their mouths, which inflate when they croak, amplifying the sounds.
The skin of amphibians is bare and moist, covered with mucus.
Body coloring helps protect against enemies.
2. Draw a drawing of the frog’s body and label its sections.

3. Consider the structure of the front and hind limbs. Sketch them.

4. Examine the frog's head. What sense organs are located on it?
see table No. 144
5. Note the structural features of the frog associated with life in water and on land.
In water: skin is bare, covered with mucus. There are nostrils on the head and eyes on the top. There are swimming membranes on the paws. The hind legs are long. Development and reproduction in water. In water it switches to skin respiration. Cold-blooded. The larva has structural features similar to fish.
On land: 2 pairs of limbs, moves. Breathes with lungs. Feeds on insects. The heart is three-chambered.
Conclusions: amphibians are the first chordates to reach land. They still have external and internal structural features that allow them to partially live in water, however, they also have progressive structural features characteristic of terrestrial animals.









